
NEET Biology Notes For Sexual Reproduction In Flowering Plants Introduction
R. Camerarius was the first to describe sexual reproduction in plants. The sexual reproductive organ in angiosperms is flower. The male sex organs are known as stamen or microsporophyll and the female sex organs are known as pistil or carpel.
NEET Biology Notes For Sexual Reproduction In Flowering Plants Stamen Or Microsporophyll: Male Sex Organ
The fertile portion of stamen is called anther. Each anther is usually made up of two lobes connected by a connective. A typical anther consists of two lobes or four microsporangia; such an anther is called bithecous. In the members of Malvaceae, arther consists of one lobe or two micro-sporangia; such an an er is called monothecous and is bisporangiate. In Arceutho- bi, the smallest dicot parasite, anther consists of only one microsporangium (monosporangiate).
reproduction flower
Development of Anther
A young anther consists of homogeneous mass of meristematic cells called primary sporogenous cells surrounded by anther wall. Primary sporogenous cells form microspore mother cells (2n) inside the microsporangium.
The anther wall consists of the following layers:
- Epidermis: It is single layered and protective in function.
- Endothecium: The cells of this layer have a-cellulosic fibrous bands arising from inner tangential wall which help in the dehiscence of anther. Fibrous bands are ab- sent in hydrophytes.
- Middle layer: The cells of this layer are ephemeral and are three-layered.
- Tapetum: This is the innermost layer of anther wall which surrounds the sporogenous tissue. Tapetal cells are nutritive. They are multinucleate and polyploid. The tapetum has two types of cells:
- Secretory or glandular: These cells secrete sporopollenin, pollen kit, and compatibility proteins. The Ubisch bodies are present in these cells, which help in the ornamentation of exine, as they have a chemical called sporopollenin deposited on them.
- Amoeboid or plasmodial or invasive: Cells undergo breakdown and entire protoplast moves in the center and nourishes microspores.
Read and Learn More NEET Biology Notes
Microsporogenesis
The formation and differentiation of microspores (pollen grains) is called microsporogenesis. In the cavity of microsporangium, the microspore mother cells divide meiotically to produce microspore tetrads or pollen tetrads.
Cytokinesis may occur either after each meiotic division (successive type) leading to the formation of isobilateral tetrad of microspores (e.g., monocots) or after both meiotic (1 and 2) divisions (simultaneous type) leading to the formation of tetrahedral tetrad of microspores (e.g., dicots). Successive type of cytokinesis is advanced type.
Tetrads are of five types: (a) tetrahedral, (b) isobilateral, (c) T-shaped, (d) linear, and (e) decussate.
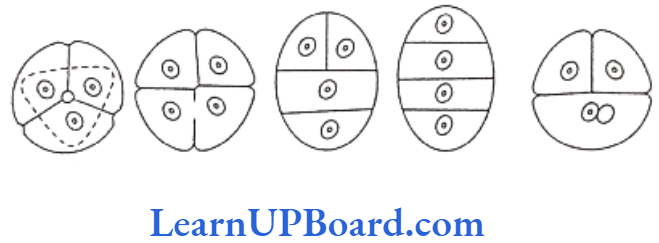
In Aristolochia elegans, all five types of tetrads are present.
Some Exceptions to Remember
- After formation, the microspores are separated from the tetrad. But in Elodea, Drosera, and Typha, the microspores do not separate from each other. Thus, com- pound pollen grains are formed.
- In Asclepiadaceae (Calotropis) and Orchidaceae, all microspores in a sporangium adhere together in a single mass known as pollinium.
- In Calotropis, the pollinia of adjacent anthers of different stamens are attached by thread-like caudicles to a sticky disc called corpusculum. The whole structure is called translator.
- In Cyperaceae, out of four microspores formed, three are degenerate. So, ultimately one microspore mother cell (2M) produces only one microspore or pollen grain.
- Sometimes, more than four pollen grains are produced from one microspore mother cell. It is called polyspory. For example, Cuscuta reflexa.
” reproduction in angiosperms”
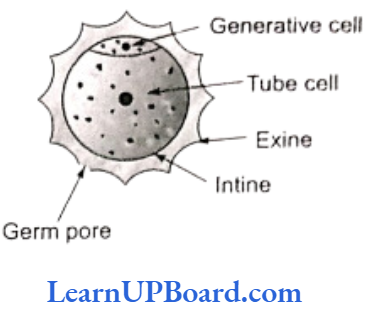
Structure of Microspore or Pollen Grain
The cell wall of microspore (sporodern) consists of two layers. The outer layer is exine and the inner layer is intine. Exine is made up of sporopollenin and intine is pectocellulosic in nature. Sporopollenin, a fatty substance, is resistant to physical and biological decomposition. So pollen wall is preserved for long periods in fossil deposits. Pollen kit is a sticky layer found on the outside of mature pollen grains of many insect pollinated species. The material of pollen kit is contributed by the tapetal cells. It acts as an insect attractant.
Exine has proteins for enzymatic and compatibility reactions. It is two-layered:
- Ektexine: Highly sculptured and is differentiated into tectum, baculum, and foot layer
- Endexine: Not sculptured
Some weak points are present on the exine. These are called germ pores.
Pollen grains can be monocolpate (having one germ pore called germinal furrow, e.g., monocots), bicolpate (two germ pores), and tricolpate (three germ pores, e.g., dicots).
The study of pollen grains is called palynology (term given by Hyde and Williams).
Male Gametophyte
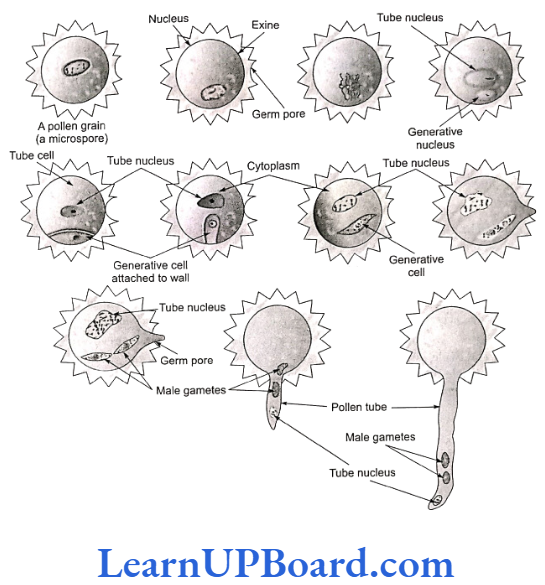
Microspore is the first cell of male gametophyte. Its germination starts in situ (in the mother place). It can be best defined as partially developed male gametophyte.
Microspore divides mitotically into large tube cell and small generative cell. At this two-celled stage (in some cases at three-celled stage), pollination takes place. Further development of male gametophyte takes place on the stigma.
Pollen grain expands by absorbing liquid from the moist surface of stigma. Stigma provides boron, sugar, amino acids, etc. The exine bursts and the intine comes out in the form of pollen tube.
Pollen grains are either monosiphonous (with one pollen tube) or polysiphonous (with more than one pollen tubes). For example, members of Cucurbitaceae and Malvaceae. Pollen tube was first observed by G. B. Amici (1824) in Portulaca.
The generative nucleus divides mitotically to form two male gametes called sperms. The male gametes are non-motile and amoeboid. They are slightly unequal in size.
The function of the pollen tube is to carry sperm. In the pollen tube, the tube nucleus enters first (it is a vestigial structure) and soon disintegrates. The growth of the pollen tube is apical and the entire cytoplasm of pollen grain is confined to the tip of the pollen tube.
NEET Biology Notes For Sexual Reproduction In Flowering Plants Carpel Or Megasporophyll: Female Sex Organ
A carpel consists of ovary, style, and stigma. Ovary contains ovules.
Structure of Ovule or Integumented Megasporangium
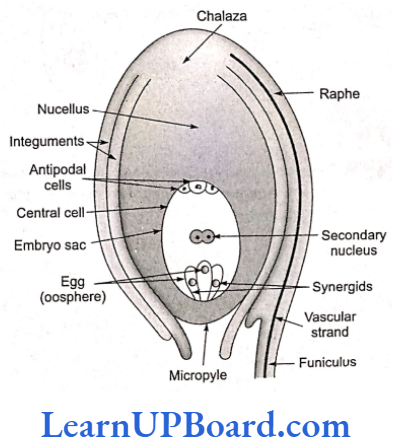
Ovule is an outgrowth of placenta. Each ovule is attached to its placenta by a stalk known as funicle. The point of attachment of the funicle with the main body of the ovule is called bilum. Sometimes funicle gets fused with the body of the ovule along one side and forms a ridge known as raphe. The basal region of the ovule is known as chalaza.
The main body of an ovule is called nucellus (called mega- sporangium) which consists of mass of parenchymatous tissue. On the basis of the development of nucellus, ovules are of two types:
- Crassinucellate: The nucellus is well developed, e.g., polypetalae.
- Tenuinucellate: The nucellus is poorly developed, e.g., gamopetalae.
The nucellus is invested all around by one or two ring-like coverings called integuments except at the apex where a small passage is formed known as microplyle. On the basis of the number of integuments, ovules are of the following types:
- Unitegmic: These are ovules with one integument, e.g., members of gamopetalae and gymnosperms.
- Bitegmic: These are ovules with two integuments, e.g., members of polypetalae and monocots.
- Ategmic: These are ovules without integument, e.g., Santalum, Loranthus (parasites), and Viscum.
The third integument in the form of aril develops from the base of ovule or funicle in many plants, e.g., litchi and Inga dulce. In litchi and Inga dulce, aril is fleshy and edible.
Types of Ovules
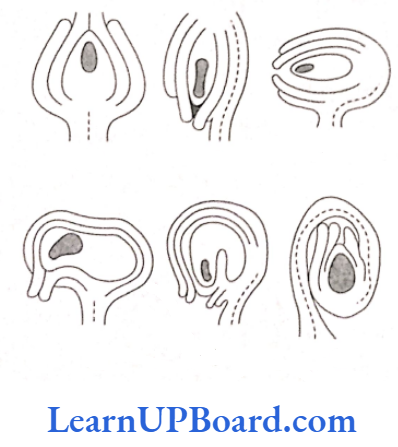
- Orthotropous: The micropyle, chalaza, and funicle are in straight line. This is the most primitive type of ovule. For example, piper, polygonum, and cycas.
- Anatropous: The ovule turns at 180° angle. Thus, it is inverted ovule. Micropyle lies close to hilum or at the side of hilum. For example, it is found in 82% angiosperm families.
- Hemianatropous: The ovule turns at 90° angle upon the funicle or the body of ovule and is at right angle to the funicle. For example, Ranunculus.
- Campylotropous: The ovule is curved more or less at right angle to the funicle. The micropylar end is bent down slightly, for example, in the members of Leguminosae and Cruciferae.
- Amphitropous: The ovule as well as embryo sac is curved like horseshoe. For example, Lemna, poppy, and Alisma.
- Circinotropous: The ovule turns at more than 360° angle and, so, the funicle becomes coiled around the ovule. For example, Opuntia (Cactacea).
reproduction of flowering plants
Megasporogenesis
Any cell of nucellus towards the micropylar end is differentiated from the other cells. This cell is called the megaspore mother cell (MMC). It divides meiotically to produce linear megaspore tetrad. In majority of angiosperms, the chalazal megaspore is functional and the other three megaspores degenerate.
Female Gametophyte of Embryo Sac
P. Maheshwari classified embryo sac, on the basis of number of megaspore nuclei participating in its formation, into the following:
- Monosporic embryo sac: Only one megaspore nucleus forms embryo sac. For example, Polygonum and Oenothera.
- Bisporic embryo sac: Two megaspore nuclei take part in the development of embryo sac. For example, Allium and Endymion.
- Tetrasporic embryo sac: All four megaspore nuclei take part in the development of embryo sac. For example, Adoxa, Plumbago, Drusa, Fritillaria, Penaea, Plumbagella, and Peperomia.
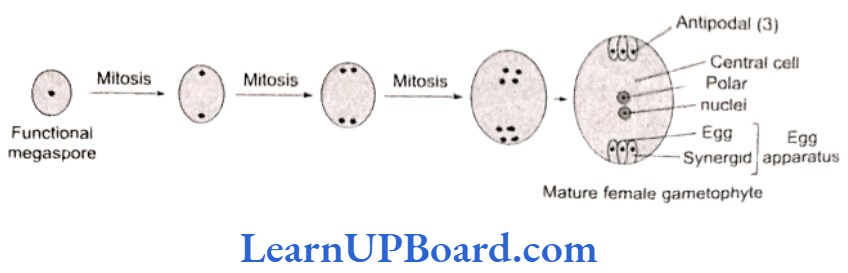
Development of Monosporic Embryo Sac (Polygonum Type)
The normal type of embryo sac development has been studied in Polygonum by E. A. Strasburger. Since this embryo sac develops from one megaspore, it is monosporic embryo sac. It develops from chalazal functional megaspore (fourth from micropyle). The nucleus of functional megaspore divides by three mitotic divisions to form eight nuclei.
- Three cells at the micropylar end form egg apparatus. One is egg cell (n) and two are synergids (n) or cooperative cells.
- Three cells at the chalazal end form antipodals (n) or vegetative cells of gametophyte.
- Two nuclei (one from each pole) in the center are called polar nuclei (n).
flowering plants reproduction
As the embryo sac matures, these polar nuclei get fused to form a secondary nucleus (2n) or definitive nucleus. An embryo sac is seven-celled and eight-nucleated structure.
NEET Biology Notes For Sexual Reproduction In Flowering Plants Pollination
The transfer of pollen grains from the anther of a flower to the stigma of the same or different flower of the same species is called pollination. It is of two types:
- Self-pollination or autogamy: When pollen grains are transferred from the anther to the stigma of the same flower, the process is called self-pollination or autogamy.
- Bisexuality: The flower should be bisexual or hermaphrodite. For example, Cartharanthus.
- Homogamy: Male and female reproductive parts in a bisexual flower mature at the same time. For example, Mirabilis.
- Cleistogamy: Sometimes bisexual flowers remain closed and never open. Such flowers are known as cleistogamous. For example, Commelina benghalensis, Viola, Oxalis, and Arachis.
- Bud pollination: Self-pollination occurs in the bud stage before the opening of flowers. For ex- ample, Pisum, wheat, and rice.
- Cross-pollination or allogamy: When pollen grains are transferred from the anther to the stigma of the flower of another plant of the same or different species, the process is called cross-pollination or allogamy. It is of two types:
- Geitonogamy: Pollination taking place between two flowers of the same plant (genetically self- pollination but ecologically cross-pollination)
- Xenogamy: Pollination taking place between two flowers of different plants (genetically and ecologically cross-pollination)
Contrivances for Cross-Pollination
- Unisexuality or dicliny: It is the formation of unisexual flowers. In unisexual flowers, allogamy becomes obligatory. Unisexuality can be seen in monoecious plants, e.g., maize and Vallisneria.
- Dichogamy: In bisexual flowers, the two sexes mature at different timing. When anthers mature first, it is called protandry. For example, sunflower and cotton. When gynoecium matures first, it is called protogyny. For example, Vallisneria.
- Heterostyly: Flowers are dimorphic with regard to the length of style and, thus, facilitate cross-pollination. For example, Primula (primrose) jasminum.
- Herkogamy: It is the presence of natural and physical barrier between androecium and gynoecium. For example, Gloriosa and Salvia.
- Self-sterility or incompatibility: Due to physiological or genetic reasons, the pollen fails to germinate on its own stigma. For example, tobacco.
Agencies for Cross-Pollination
- Entomophily (pollination by insects): 80% of in- sects’ pollination occurs by bees (chief pollination). All flowers pollinated by bees are brightly colored, have a sweet smell, and produce nectar. Entomophil- ous flowers produce a small amount of pollen which has a spinous and sticky exine due to the presence of pollen kit. The stigmas of such flowers are long, rough, and sticky. Salvia is an excellent example of insect pollination, which occurs by lever or turn-pipe mechanism. Calotropis exhibits translator mechanism. Aristolochia shows pitfall or flytrap mechanism. Moth-pollinated plants are white flowered and fragrant. Pollination in orchid (Ophrys speculum) occurs by wasp (pseudocopulation mechanism). In Ficus carica, pollination occurs by an insect Blastophaga (trapdoor mechanism). Yucca is pollinated by Pronuba yuccasella (obligate relation between two).
- Anemophily (pollination by wind): It is a non-directional and wasteful process. Windpollinated flowers produce a large number of pollen grains to compensate for the wastage. Female flowers have large feathery brush like stigmas to catch the pollen grains. Anemophilous flowers are small and inconspicuous with long and versatile stamens. Pollen grains are dry and powdery and are produced in large numbers. For example, sugarcane, maize, wheat, bamboo, Pinus (winged pollen), and papaya.
- Hydrophily (pollination by water): All aquatic plants are not hydrophilous.
Some hydrophytes are anemophilous, such as Potamogeton and Myriophyllum, or entomophilous, such as Alisma and lotus. Hydrophilous plants may be pollinated inside the water (hypohydrophily) as in Zostera and Ceratophyllum or outside the water (epihydrophily) as in Vallisneria (tape grass and ribbon weed).
” reproduction in flowering plant”
- Ornithophily (pollination by birds): Flowers are brightly colored but are odorless and produce plenty of nectar and large quantities of pollen. For example, Bombax, Callistemon, Strelitzia, and Erythrina.
- Chiropterophily (pollination by bats): Bats pollinate the flowers of tropical regions. For example, Anthocephalus, Kigelia, and Adansonia.
- Malacophily (pollination by snails): For example, arum lillies, Arisaema, and Lemna.
- Ophiophily (pollination by snakes): For example, Santalum and Michelia.
Entry of Pollen Tube into Ovule
A pollen tube mostly enters into an ovule through the micropyle; it is called porogamy, as seen in most of the flowering plants. In some plants, such as Casuarina, the pollen tube enters the ovule through chalaza; it is called chalazogamy. Sometimes it enters through integuments; it is called mesogamy. For example, Cucurbita.
NEET Biology Notes For Sexual Reproduction In Flowering Plants Double Fertilization
Pollen tube enters one of the synergids and bursts releasing two male gametes. One male gamete fuses with the egg to form dip- loid zygote. The fusion is called syngamy or generative fertili- zation. It was discovered by Strasburger.
Male gamete (n)+ Egg (n) → Zygote (2n)
The other male gamete fuses with the secondary nucleus (2n after the fusion of polar nuclei) to form primary endosperm nucleus (3n). This fusion is also called triple fusion because three nuclei take part in this. It is also known as vegetative fertilization or pseudofertilization or trophomixis.
Male gamete (n)+ Secondary nucleus (2n) → Primary endosperm nucleus (3)
Both syngamy and triple fusion are called double fertilization. As the fusion in the embryo sac occurs twice, this is called double fertilization. Triple fusion and double fertilization were discovered by S. G. Nawaschin and Guignard in Lilium and Fritillaria, respectively. Double fertilization occurs in angiosperms only. In all, five nuclei are involved in double fertilization.
Endosperm
An endosperm is a product of triple fusion and develops from the central cell of embryo sac. It is generally a triploid tis- sue. This endosperm is the nutritive tissue for the developing embryo. It is absent in families such as Orchidaceae and Podostemaceae, and trapaceae.
Nature of Endosperm
- Cells are isodiametic and polyploid.
- Starch endosperm in cereals.
- Proteinaceous endoperm (aleurone layer) in cereals.
- Oily endosperm in castor and coconut.
- Cellulosic endosperm in ivory palm (hard endosperm).
- Hemicellulosic endosperm in date palm.
Types of Endosperms
- Nuclear endosperm: Primary endosperm nucleus of the central cell divides without wall formation (free nuclear division). It is the most common type of endosperm. For example, cotton, maize, Capsella, and coconut (milk).
- Cellular endosperm: Primary endosperm nucleus divides many times and each division is followed by wall formation. For example, Petunia, Utricularia, and coconut (copra).
- Helobial endosperm: It is intermediate between nuclear and cellular types. For example, members of order Helobiales (monocot).
NEET Biology Notes For Sexual Reproduction In Flowering Plants Embryo
The study of the development of embryo is called embryogeny.
Development of Embryo in Dicots
The normal type of dicot embryo development has been studied in shepherd’s purse (Capsella bursa-pastoris) which belongs to family Cruciferae. This is called as crucifer or onagrad type of embryo development.
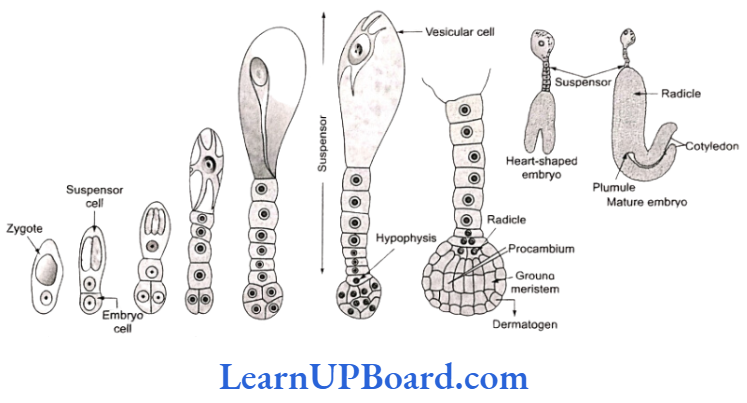
The development of embryo is endoscopic. The zygot (oospore) divides into two unequal cells a larger sus- pensor cell towards micropyle and a smaller embryonal cell (or terminal cell) towards the antipodal region. The suspensor cell undergoes transverse divisions forming 6-10 celled long sus- pensor. The first cell of the suspensor (towards embryo cell) is known as hypophysis. It forms the radicle tip.
The embryonal cell divides twice vertically and once trans- versely to produce a two-tiered eight-celled embryo. The epibasal tier forms two cotyledons and a plumule while the hypobasal tier produces only hypocotyl and most of the radicle. For this, the octant embryo undergoes periclinal divisions producing protoderm, procambium, and ground meristem. It is initially globular, but with the growth of cotyledons, it becomes heart-shaped and then assumes the typical shape. For example, Capsella bursapastoris. In orchids, Orobanche, and Utricularia, the embryo does not show the distinction of plumble, cotyle- dons, and radicle.
Development of Embryo in Monocot
The normal type of monocot embryo development has been studied in Luzula forsteri and is called Sagittariatype.
The early developments of dicot and monocot embryos are similar up to the octant stage. Later on differentiation starts. Suspensor is single celled.
The zygote of oospore divides transversely producing a vesicular suspensor cell towards the micropylar end and an embryo cell towards the chalazal end. The embryo cell divides transversely again into a terminal and a middle cell. The terminal cell divides vertically and transversely into globular embryo. It forms a massive cotyledon and a plumule.
The growth of cotyledon pushes the plumule to one side. The remains of the second cotyledon occur in some grasses. It is called epiblast. The single cotyledon of monocots is called scutellum. It is shield-shaped and appears terminal. The middle cell gives rise to hypocotyl and radicle. It may add a few cells to the suspensor. Both radicle and plumule develop covering sheaths called coleorrhiza and coleptile, respectively.
Incompatibility
Incompatibility is the inability of certain gametes, even from genetically similar plant species, to fuse with each other. This is also called intraspecific incompatibility, self-sterility, and self- incompatibility. It involves many complex mechanisms associated with the interaction of pollen and stigmatic tissues. It is of two types:
- Sporophytic incompatibility: It is the incompatibility due to the genotype of sporophytic/stigmatic tissues.
- Gametophytic incompatibility: It is the incompatibility due to the genotype of the pollen. This may be due to the prevention of pollen germination, deorientation of pollen tube, or even failure of nuclear fusion. It is controlled by genes with multiple alleles (S-allele). A plant carries two such alleles, e.g., S1, S2, S2, S3, S1, S3, S2, S4, S3, S5, etc. A pollen carries only one allele. If it happens to be one of the two alleles of pistil, the pollen fails to form pollen tube.
Apomixis and Polyembryony
Apomixis is the formation of new individuals directly through asexual reproduction without involving the formation and fusion of gametes. It is of two types: (a) agamospermy and (b) vegetative propagation.
NEET Biology Notes For Sexual Reproduction In Flowering Plants Agamospermy
In this type of asexual reproduction, the embryo is formed by a process in which normal meiosis and syngamy have been eliminated. This type of apomixis occurs within the seed.
Types of Agamospermy
- Adventitive embryony (sporophytic budding): The embryo arises from diploid sporophytic cells such as nucellus or integuments (other than egg). Examples are Citrus and Opuntia.
- Recurrent agamospermy: In this method, a diploid embryo sac is formed from MMC which has a diploid egg or oosphere. The diploid egg grows parthenogenetically into diploid embryo. Examples are apple, pear, and Allium.
Diploid embryo sac can develop directly either from the diploid MMC (diplospory) or the diploid nucellai cell (apospory). - Non-recurrent agamospermy: The embryo develops parthenogenetically from the haploid egg. Example is banana.
Polyembryony is the phenomenon of having more than one embryo. There may be more than one egg cell in an embryo sac or more than one embryo sac in an ovule. All the eggs may get fertilized. Synergids and antipodal cells may also form embryos. The occurrence of polyembryony due to the fertilization of more than one egg is called simple polyembryony.
The formation of extra embryos through sporophytic budding is called adventive polyembryony. Polyembryony is quite common in onion, groundnut, mango, lemon, and orange. In some of these cases, a stimulus of pollination may be required. Citrus seed has 2-40 embryos-one normal and the rest adventitive mostly nucellar.
NEET Biology Notes For Sexual Reproduction In Flowering Plants Assertion-Reasoning Questions
In the following questions, a statement of Assertion (A) is followed by a statement of Reason (R).
- If both Assertion and Reason are true and the reason is the correct explanation of the assertion, then mark (1).
- If both Assertion and Reason are true but the reason is not the correct explanation of the assertion, then mark (2).
- If Assertion is true but Reason is false, then mark (3).
- If both Assertion and Reason are false, then mark (4).
Question 1. Assertion: All the pollen grains of a microsporangium are held together and form rollinium.
Reason: Pollinium is very suitable for anemophily.
Answer. 3. If Assertion is true but Reason is false, them mark (3).
Question 2. Assertion: An endosperm represents the triploid condition.
Reason: It is formed due to the fusion of triploid nuclei.
Answer. 3. If Assertion is true but Reason is false, them mark (3).
Question 3. Assertion: Allele of pollens happens to be one of the two alleles of pistil; the pollen fails to form pollen tube.
Reason: The incompatibility is due to the genotype of the pollen.
Answer. 1. If both Assertion and Reason are true and the reason is the correct explanation of the assertion, then mark (1).
Question 4. Assertion: The pollen grains of dicot are tricolpate.
Reason: It has three distinct lens shaped apertures,
Answer. 1. If both Assertion and Reason are true and the reason is the correct explanation of the assertion, then mark (1).
Question 5. Assertion: The main body of an ovule is called nucellus.
Reason: The nucellus is well-developed in polypetalac.
Answer. 2. If both Assertion and Reason are true but the reason is not the correct explanation of the assertion, then mark (2).